

What is Africa?
Part 1: Africa “proper”

Written by: Timothy Crowe
My goals are to succinctly address past and current answers to this ‘multi-answered’ questions.
Geography and Geomorphology
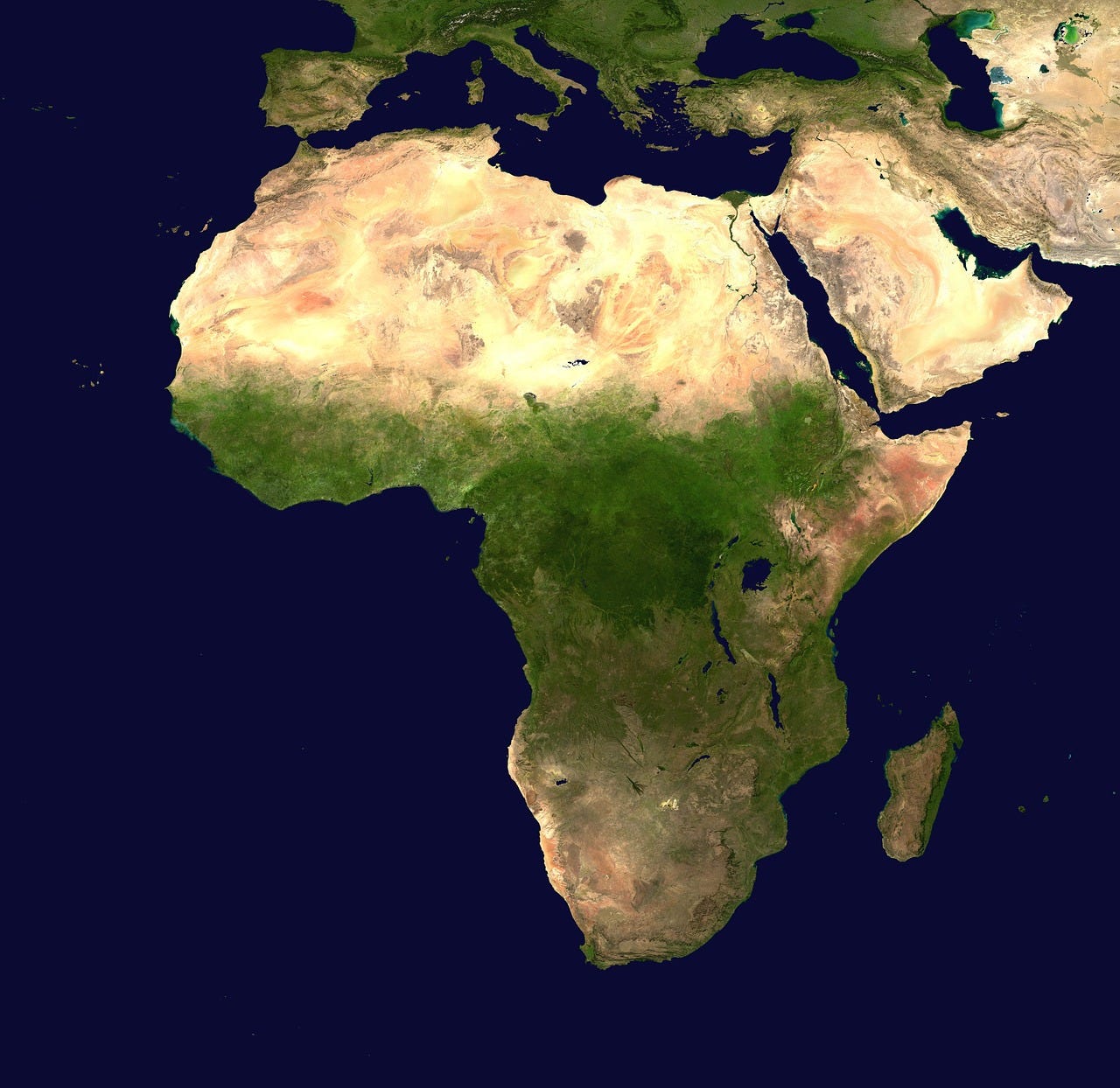
Africa is the second largest continent (ca 30 million km2,), 31% of which encompasses the Sahara - Earth’s largest hot desert. It is divided roughly in half by the equator and surrounded by sea, except where the Isthmus of Suez joins it to Asia. It became isolated from the other large ‘chunks’ of the southern super-continent Gondwana by tracts of water at about 80 million years ago (mya) and ‘collided’ with Eurasia about 20 mya. Geomorphologically, Africa is ‘fractured’ into two tectonic plates which have been diverging from one another since about 50 mya. The Nubian Plate encompasses all of northern, central and southwestern Africa. The Somali Plate covers much of eastern Africa. The boundary between these two Plates extends from the Gulf of Aden in the north through the East African Rift Valley south to South Africa, and the Plates are moving apart at an average rate of about 2.5 cm per year.
Biotic turnover
About 65.5 mya, during the K–Pg extinction event at the end of the Cretaceous period, there was a sudden, mass global extinction of ca 80% of vertebrate species. Thereafter, there was an ‘explosive’ evolutionary radiation within Africa. However, especially for birds and mammals, most of this evolution was confined, biogeographically, to the Afrotropical Realm (one of Earth's eight) which includes only Africa south of the Sahara. Perhaps the best mammalian example of a Afrotropical radiation is the Afrotheria, a basal group of morphologically highly dissimilar placental mammals whose discovery in the 1990s was based largely on DNA evidence. It includes the golden ‘moles’, elephant shrews, tenrecs, aardvarks, hyraxes, elephants and sirenians (dugongs and manatees).
‘Birdwise’, there are the African endemic guineafowls (Numididae), chicken-like gamebirds with genera confined to primary equatorial forests (Agelastes spp.), secondary/riverine forests (Guttera spp.), savannas (Numida) and deserts (Acryllium).
The flora and fauna of northern, supra-Saharan Africa is qualitatively distinct and has strong evolutionary and biogeographical connections with the biota of the Palaearctic Realm which stretches across Eurasia.
With regard to the Sahara, about 8500 years BCE, after a long arid period during the last ice age, it began to experience heavy monsoons. The ‘palaeo-Sahara’ became a grassy savanna, full of edible plants and animals. Dark-skinned prehistoric settlers moved in from the Nile valley and developed cattle pastoralism and the hunting of wildlife. However, by about 5300 BCE, the monsoons began to dry up, ‘recreating’ a massive desert, and Saharan peoples began to cluster back around the Nile again. The return of full desert conditions at about 3500 BCE coincided with the initial stages of pharaonic civilization in the Nile valley.
Afrotropical biogeography
With respect to mammals and non-aquatic birds, within sub-Saharan Africa, there are five well-defined - but evolutionarily spatio-temporally dynamic - biogeographical Regions that may be sub-divided into Provinces and Districts delineated by valleys, mountains/ranges and sharp climatic transitions (see here and here). The Equatorial Forest centres on the Congo River Basin and may be sundered into island-like Eastern, Western and Far-western ‘refugia’ which exist during macro-climatic dry periods. The Northern Savannas may be cleaved into Eastern and Western districts by a sometimes mega-Lake Chad. Also during macro-climatic dry periods, the desertic Northeastern ‘Horn’ of Africa is connected – by an Arid ‘Corridor’ - to the Southwestern Deserts centred on Namibia, Botswana and the South African Karoo. The biogeographic ‘conduit’ connecting the Afrotropics and the Palaearctic is in northeastern Africa along the 7000 km Nile River drainage basin.
Stylized African biogeographical zones: Palaearctic North Arica (grey); Deserts (yellow); Savannas (brown) split near the Equator – northern sometimes sundered by mega-Lake Chad (blue) – southern sometimes split north-to-south by Arid ‘Corridor’; Equatorial Forest (green) – during dry periods reduced to three ‘refugia’ (outlined in black) separated by savanna ‘corridors’ (light green)
Depending on changes in macro-climate, forests, savannas and deserts expand and contract geographically, creating and undermining island-like entities within which groups of ‘higher’ African vertebrates have speciated - or ‘attempted’ to speciate. This process is well illustrated in gamebirds [guineafowls (Numididae), francolins (Francolinus, Ortygornis, Afrocolinus, Peliperdix, Scleroptila and Campocolinus spp.) and spurfowls (Pternistis spp.) and Primates [e.g. Baboons (Papio and Theropithecus spp. – see here, here, here - and hominins, including Modern Humans.
Timothy Crowe is an emeritus professor of evolutionary and conservation biology.